
| ← トップページに戻る | |
|
トピックス
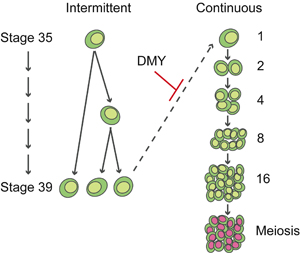
性分化風々譚 −生殖細胞が関わる性分化− 性の多様なところと保守的なところ 多くの脊椎動物ではまず性的未分化の生殖腺原基で性決定が起こり、その後の性分化で卵巣あるいは精巣が形成される。人間社会で生活していると、生まれつき性染色体によって性が決まる(Y染色体の精巣決定遺伝子SRYがあると雄)のが当たり前と感じるかも知れないが、生き物全体では決してそうではない。は虫類のように温度などの環境要因によって性が決まる動物もいる。 雌雄の成立の仕方(性分化)も様々で、ある種の魚類のように雄から雌へと(精巣から卵巣へと)転換する種もあれば、その逆に転換する種もある。卵巣・精巣両方をもつ種も知られている。ヒトなど哺乳類の一方向的様式は性分化様式の1類型でしかないことが判る。このような性決定・性分化の多様性は、それぞれの生物種が最適な繁殖戦略の達成するために進化の過程でもたらされたと理解することは可能である。「最適なタイミングで、最適な数の、性的に成熟した雌雄が存在すること」が重要であり、生態学の言葉を借りれば「如何に繁殖コストを下げて次世代を作るか」、生殖生物学で言えば「如何に配偶子形成を行ないと有性生殖を達成するか」、で多様性が産み出されるとも説明される。 それでは性決定分化の多様な現象の部分では、遺伝子・細胞レベルのメカニズムとしていったい何がおきているのであろうか?生物の発生現象を見ていると、基本現象と思われるところでは頑なまでに保守的な機構を保持している。前後・背腹軸決定とhox遺伝子との関係然り。ここではハエであろうとヒトであろうとその仕組みに区別はない。「雌雄という二型がある」ことに関しても生き物はきわめて保守的である。雌雄とは「卵と精子と言う2種類の配偶子がある」に対応した現象であり、「卵巣と精巣という2つの器官」を基礎とした現象である。一方で性分化の多様性そのものが繁殖戦略の要として保存されているならば、共通の卵巣・精巣形成から多様性を産み出すための何らかの基本原理があるのだろうか?何らか生き物共通の遺伝子レベル、細胞レベルの仕掛けがあるのであろうか? 特定領域研究での研究ことはじめ 特定領域研究での4年に渡る公募研究では、メダカを用いて生殖腺を構成する様々な細胞を特定し、その細胞系譜解析を行ない、それら細胞の性分化における機能を生殖腺形成や性分化がおかしくなる突然変異体と組み合わせ、生殖腺形成と性分化過程の解析を試みてきた。性決定分化の研究では先行研究としてマウスでの研究がある。遺伝子破壊実験や組織培養の技術、系統間の遺伝学的解析を活用し、常に性決定分化研究の先導役となってきた。 しかし私にはどのように性が決まって卵巣・精巣へと分化するのか、理解できないところが多々あった。とりわけ、個体の中で細胞レベルでの性分化がどう進むのか、またそれがどのように性特異的組織の構築に関与するのか、具体的にイメージしにくい部分が多々あった(それはメダカを用いた今でも同じではあるが、、、)。マウスにおいても(勿論ヒトにおいても)、生殖腺にどのような細胞が存在するのか、組織学的に見える多くの細胞については実はよく判っていないと思う。性分化過程では細胞に障害があると生殖腺の形成不全を引き起こすだけでなく性転換を引き起こすことがある。このことは性分化過程では細胞レベルの適切な性分化が必要で、しかもその性分化状態を細胞が積極的に維持していることを示唆している。性分化現象を理解しようと思うとこの細胞レベルの事象を知ることが「私にとって」性分化現象を一番「諒解」させてくれるだろうと思えたし、またメダカを用いるならばこの細胞レベルの現象を見てみたいと思い、華やかな分子機構解析ではなく、細胞系譜解析を研究の柱において地道に研究を進めることにした。 体細胞の性によって制御される生殖細胞の分裂様式 メダカの胚や幼魚は基本的に透明であり、ある細胞に蛍光蛋白質を特異的に発現させることでその細胞を個体内で特定、その細胞の系譜を解析することが可能である。我々は、生殖腺の主要な構成細胞ともみなすことのできる生殖細胞を可視化して、生殖細胞の挙動をまず解析することにした。 メダカは体節形成中期の受精後3日目頃(stage 23 以降)、側板中胚葉後端部から分化した異なる2種の生殖腺体細胞と始原生殖細胞があわさり、後腸背側に移動し、受精後4日目ごろに(stage 30 前後)生殖腺原基を形成する(Nakamura et al., 2006)。この後、雌で生殖細胞が増殖し、受精後7日-10日の孵化の時期になると明らかに雌の生殖腺が大きくなっている。この性分化初期の過程は生体内での観察が比較的しやすかったため、生殖細胞の1部のみを蛍光でラベルして、その分裂の様子を観察した。 すると生殖細胞は、雌雄両方の生殖腺で間欠的な分裂を行ない、その娘細胞は直ちにお互い分離する分裂様式が観察された。これに加えて雌では、連続的に数回分裂し、娘細胞も離れないでかたまって生殖細胞シストを形成する分裂様式が見られた。前者の間欠的分裂をタイプI 型、後者の連続分裂をタイプII 型分裂と呼ぶことにした(図1)。一方、生殖細胞が生殖腺形成初期は存在するがやがて失われて不稔となる突然変異体メダカ zenzai が単離されていたので、この生殖細胞を観察したところ、タイプI型を行なう生殖細胞に二核化が生じて最終的に全ての生殖細胞が失われることが判明した。タイプII型分裂を行なう生殖細胞は正常でしかも減数分裂マーカーを発現することから、タイプII型分裂は配偶子形成過程の分裂、タイプI型分裂は生殖細胞維持の分裂と結論された(Saito et al., 2007)。メダカ生殖腺の雄化は体細胞側で性決定遺伝子が発現して行なわれることから、この生殖細胞分裂の性的二型は体細胞で制御されると考えられ、この分裂様式の違いがメダカ雌雄の生殖腺の大きさの違いをもたらしていると結論された(図2)。生殖細胞自体も異なる分化段階にあることが、生殖顆粒の細胞内局在変化からも示唆されている(Aoki et al., 2008)。
性分化における生殖細胞の役割 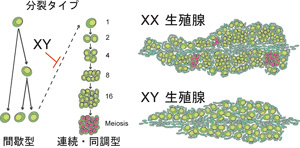 それでは生殖細胞は性分化過程に関与していないのか? その結果、以前の報告とは全く異なり、生殖細胞がないと生殖腺はチューブ状の形態を示すことが判明した。このことは生殖細胞が、体細胞の構成する性特異的な構造の形成に必須であることを示している。さらに面白いことに、生殖細胞が欠損したすべてのメダカで第二次性徴は完全に雄化、すなわち、メダカの遺伝的性決定システムとは独立して遺伝的雌(XX)のメダカも雄化を示していた。実際、このメダカの性ステロイドの量を計測すると、エストロゲンの量は検出限界以下でアンドロゲン量の増加が見られた。発生過程の詳細な解析では、生殖腺の体細胞も性分化開始直後はひとたび雌になるものの、直ちに雌特異的遺伝子発現し、雄型生殖腺体細胞へと分化としていた(Kurokawa et al., 2007)。 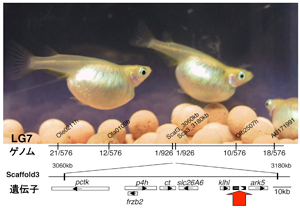 この結果は生殖細胞がないときの性分化を示しているが、それならば生殖細胞が多くなるとどうなるか?我々は生殖細胞が過形成される突然変異体メダカhoteiを単離していたのでこのメダカを解析した(図4)。生殖細胞がないメダカとは反対に、遺伝的雄(XY)の約半数が雌型の第二次性徴を示し、それらは卵巣様生殖腺を有していた。これらの中には若いうちは雌として妊性を示す個体も散見された。この突然変異体の原因遺伝子はミュラー管阻害因子のII型受容体(amhrII)であることがポジショナルクローニングの結果判明し(図4)、その発現は、リガンドamhとともに生殖細胞ではなく体細胞(支持細胞)で検出された(Morinaga et al., 2007)。このことはAMH系シグナルがオートクライン的に支持細胞で働き、可視化解析で見られたタイプI型やタイプII型の生殖細胞分裂様式の制御に関与していると推察された。 突然変異体hoteiではその支持細胞の障害の結果、生殖細胞が増殖すると解釈される。そこで突然変異体hotei の生殖細胞をなくすこと行なってみたところ、遺伝的雄個体が雌化する性転換現象は全く見られなかった。このことからhotei に見られる雌化は、支持細胞の障害による体細胞自律的(直接的)な出来事ではなく生殖細胞を介していると示唆され、生殖細胞が雌化に必須であるという生殖細胞欠損メダカと現象的にも符合する。 生殖細胞と体細胞との性的な?関係 生殖細胞がないメダカは雄化し、生殖細胞が過剰に存在する突然変異体hoteiでは雌化する。一連の研究結果、生殖細胞が生殖腺を雌へと導く能力(canalization to feminization)があり、生殖腺体細胞自身は自律的(生殖細胞非存在という意味)に雄化の能力 (male predisposition)をもつことは今回明らかとなった。しかし、この2つの性転換現象がどのような機構に基づいて起きているのか、同じ細胞相互作用の表裏の関係にあるのかあるいは全く別の機構であるのか、さらには正常な性分化機構の雌雄形成とどのように対応するのか、解決すべき問題は山積みである。 面白いことに、マウスにおいても卵巣中の生殖細胞がなくなると精巣様構造が出現し、そこには雄特異的遺伝子が発現することも報告されている。また、ゼブラフィッシュでは全ての個体に最初卵巣が形成されるが、その後生殖細胞の数が減少した卵巣あるいは生殖細胞の数が増殖しなかった卵巣が生殖細胞を失いつつ精巣へと分化を開始することが報告された。メダカでも極端に生殖腺中の生殖細胞が少なくなると遺伝的雌(XX)の性が逆転し、機能的性転換雄が出現する。これらの状況を考え合わせると、生き物(脊椎動物?)の生殖細胞と体細胞との間には、雌化と雄化の性の綱引きがあるようにも思える。 後記 始めに記したように性の決定システムと性の成立過程は多様である。メダカとマウスは同じXY型染色体性決定システムでありながら遺伝子は異なる。恐らく今後沢山の異なる性決定遺伝子が同定されるだろう。一連の結果は、この多様な遺伝的性決定システムとは別の、あるいはその基礎となる、細胞相互作用レベルの基本的な性分化機構・性の維持機構が、生き物には存在していることを現象的に示し、そこには、生殖細胞という配偶子形成を行なう細胞が関わっているという結果となった。
顧みるに、この特定領域研究が始まる時の最初のニュースレターに「性の揺らぎ」と題した雑文を寄せる機会があった。そこで「揺らぎ」と記した「性の多様性を保障するもの」が、「生殖細胞と体細胞との関係」に帰するきっかけができたのはこの4年間の公募研究での望外の喜びであった。
Aoki et al., 2008 Dev.Dyn. 237, 800-807. |
|
| ↑ このページのトップに戻る ← トップページに戻る |
| Copyright (C) Grant-in-Aid for Scientific Research on Priority Areas 'Mechanisms of Sex Differentiation' All rights reserved. |