2026.05.20
細胞膜の急速な拡大を可能にする新たなメカニズムを解明 膜タンパク質を一時的に格納する細胞膜構造が細胞運動を支える(生化学分野 池ノ内 順一 教授)
- 細胞膜が急速に膨らむ際、その根元に新たな膜陥入構造(Sub-bleb invagination; SBI)が形成されることを初めて発見
- Caveolin-1やPiezo1(※1)など、特定の曲率を好む膜タンパク質がこの構造に選択的に集積することを解明
- タンパク質の過剰蓄積が膜拡大を阻害し、細胞運動を抑制することを示し、がん細胞の浸潤・転移や免疫細胞の移動の理解につながる新たな分子基盤を提示
細胞が移動する際には、細胞膜が数秒程度で急速に広がる「ブレブ(※2)」と呼ばれる膨らみが形成されます。この現象は、免疫細胞が体内を移動する過程や、がん細胞が組織内を浸潤・転移する際にも利用される重要な細胞運動様式です。しかし、細胞膜は通常わずか2〜3%程度しか伸びることができず、このような急激な膜の拡大がどのように実現されているのかは長年の未解決問題でした。
九州大学大学院医学研究院生化学分野の池ノ内順一教授、大学院システム生命科学府博士課程学生の前川悠輝と、東京大学大学院医学系研究科分子生物学分野の水島昇教授、吉井紗織助教らの研究グループは、ブレブ形成時に細胞膜の根元で新たな膜陥入構造(Sub-bleb invagination; SBI)が形成されることを発見しました。さらに、この構造が膜の曲率に応じてCaveolin-1やPiezo1といった特定の膜タンパク質を一時的に格納する役割を持つことを明らかにしました。この仕組みにより、膜の拡張に適さない膜タンパク質が膨らんでいく膜領域から排除され、細胞膜が破綻することなく効率的に広がることが可能になります。さらに本研究では、このようなタンパク質の再配置機構が破綻すると、ブレブ形成が抑制され、細胞の移動能力が低下することも示されました。これは、膜タンパク質の空間配置が細胞運動の成立そのものに直結していることを示す重要な知見です。本研究成果は、細胞がどのようにして急速な形態変化を可能にしているのかという基本原理を明らかにするとともに、がん細胞の浸潤・転移や免疫細胞の機能など、細胞運動に依存するさまざまな生命現象の理解に新たな視点を提供するものです。将来的には、細胞運動を標的とした新たな治療戦略の開発にもつながることが期待されます。
本研究成果は米国科学アカデミー紀要(PNAS)に2026年5月20日(水)(日本時間)に公開されました。
参考図:急速に拡大する膜構造(ブレブ)の根元に形成される膜陥入構造(SBI)
研究者からひとこと
この研究では、膜曲率に依存した膜タンパク質の再配置が、膜変形そのものを成立させていることを示しました。これは、膜力学と分子配置を統合的に理解する新しい視点になると期待しています。
(大学院医学研究院 池ノ内順一)
細胞は、移動や分裂、形態変化の際に細胞膜の形を大きく変化させます。特に、免疫細胞やがん細胞は「ブレブ」と呼ばれる膜の膨らみを形成しながら移動することが知られており、この運動様式は組織内での移動や浸潤に重要な役割を果たしています。ブレブ形成では、細胞内の圧力によって細胞膜が外側へ押し出され、数秒以内に急速に拡大します。しかし、細胞膜はゴムのように自由に伸びるわけではなく、破裂せずに伸びることができるのはわずか2〜3%程度とされています。そのため、この急激な膜拡大は単純な「膜の伸び」では説明できず、追加の膜面積を供給する仕組みが必要と考えられてきました。
これまでの研究では、細胞膜の内側に存在するチューブ状の陥入構造が広がることで膜面積を供給する「膜リザーバー機構」が提唱されてきました。一方で、細胞膜には多様な膜タンパク質が存在し、それぞれが特定の曲率(膜の曲がり方)を好む性質を持っています。このようなタンパク質が、急激に形を変える膜上でどのように再配置されるのかは、ほとんど分かっていませんでした。特に、Caveolin-1やPiezo1のように内向きに曲がった膜を好むタンパク質は、外向きに大きく膨らむブレブの表面とは物理的に相性が悪いと考えられます。しかし、それらのタンパク質がどのように処理されるのかについては明らかではありませんでした。
本研究では、「細胞膜が急速に広がるとき、膜タンパク質はどのように振る舞うのか」という点に着目し、その分子機構の解明を目的として研究を行いました。
本研究では、ライブセルイメージング、三次元電子顕微鏡解析(CLEM)、および分子生物学的手法を組み合わせ、ブレブ形成時の細胞膜構造と膜タンパク質の動態を詳細に解析しました(図1)。
まず、ブレブが拡大する際、その根元に複数の細長い膜陥入構造が新たに形成されることを発見しました。三次元電子顕微鏡解析により、これらの構造は細胞膜と連続した実在の膜構造であることが確認され、研究グループはこれを「Sub-bleb invagination; SBI」と命名しました。重要なのは、この構造が既存の膜の展開によって生じるのではなく、ブレブの拡大と同時にその場で新たに形成される点です。すなわち、膜の再編成は「外向きの膨張」と「内向きの陥入」が同時に進行する、双方向的な過程であることが明らかになりました。
次に、SBIに局在するタンパク質を解析した結果、Caveolin-1やPiezo1といった正の曲率を好む膜タンパク質が選択的に集積することが分かりました。一方で、PODXLやPD-L1などの膜タンパク質はブレブ全体に均一に分布しており、膜タンパク質の分布には明確な選択性が存在することが示されました。
さらに、FRAP解析により、細胞質側タンパク質であるFBP17は速やかに交換される一方、膜タンパク質であるCaveolin-1はSBI内に比較的安定に保持されることが明らかになりました。この結果は、SBIが単なる一過性の構造ではなく、特定の膜タンパク質を一時的に隔離・保持する“貯蔵コンパートメント”として機能することを示しています。また、Piezo1の活性状態を変化させる実験により、膜曲率に対する親和性の変化に応じてSBIへの局在も変化することが示され、タンパク質の分布がその曲率選好性に依存していることが明らかになりました(図2)。
機能解析の結果、Caveolin-1やFBP17を過剰に発現させると、膜の内向きの陥入が過剰に形成され、ブレブのサイズおよび頻度が低下しました。さらに、これに伴いアメーバ様運動による細胞移動能も有意に低下することが確認されました(図3)。
これらの結果から、細胞は膜を急速に拡張する際に、「膜面積の供給」と「膜タンパク質の再配置」という二つのプロセスを同時に制御していることが明らかになりました。そして、SBIがその中核的な役割を担うことが示されました。
特に、膜拡張に適さないタンパク質をSBIに一時的に隔離することで、膜の物理的な拡張を可能にしている点は重要です。本研究は、膜変形とタンパク質配置が密接に結びついた新しい細胞膜構造の制御原理を示すものです。
本研究により、細胞膜の変形は単なる物理的な現象ではなく、膜タンパク質の配置制御と密接に連動した統合的なプロセスであることが明らかになりました。この知見は、細胞がどのようにして急速な形態変化と機能維持を両立しているのかを理解する上で重要な基盤となります。今後は、このSBI構造がさまざまな細胞種や生理的状況においてどの程度普遍的に利用されているのかを明らかにすることが課題となります。特に、免疫細胞やがん細胞など、高い運動性を持つ細胞において、その機能的意義の解明が期待されます。また、細胞運動はがんの浸潤・転移や免疫応答に直結する重要な現象であることから、本研究で明らかになった膜タンパク質の再配置機構を標的とすることで、新たな治療戦略の開発につながる可能性があります。さらに、膜の曲率とタンパク質配置の関係を人為的に制御することで、細胞の形態や運動を操作する技術の開発にもつながると期待されます。
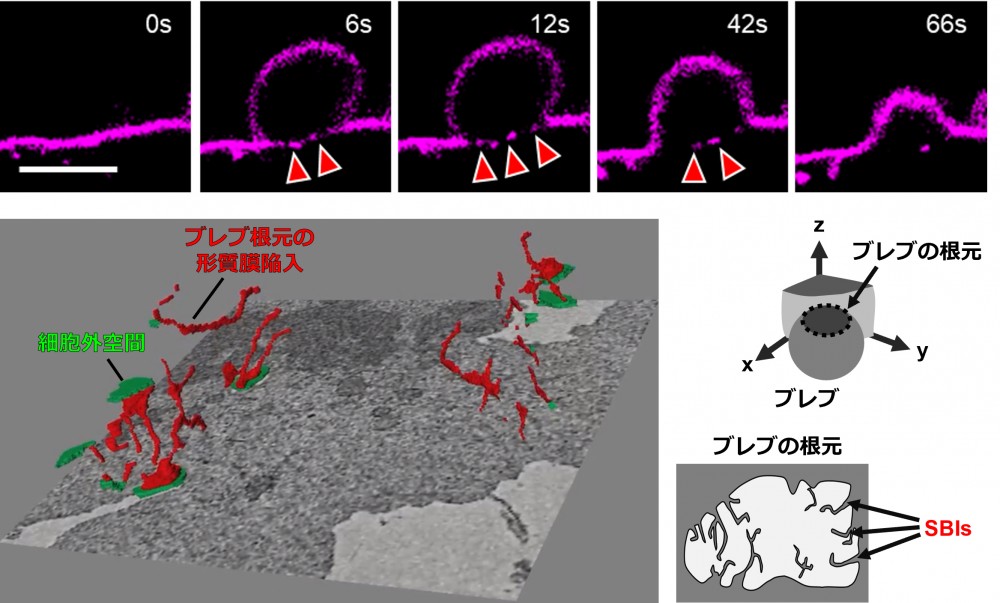
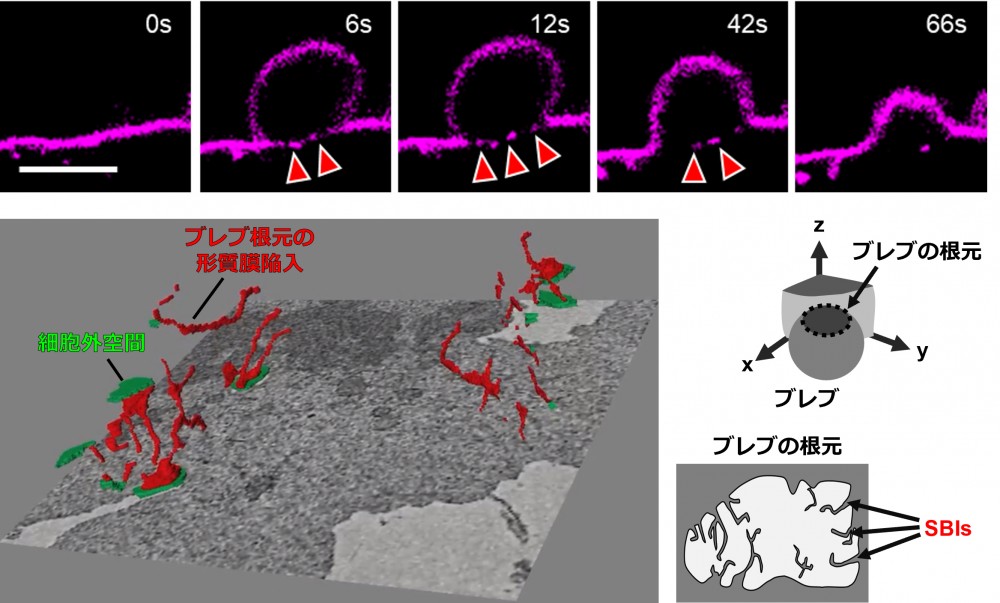
上段は、細胞膜をCellMaskで可視化した細胞において、ブレブが形成される過程をライブセルイメージングで観察したものです。ブレブは細胞膜が数秒のうちに外側へ膨らむ現象であり、細胞運動に重要な役割を果たします。矢印で示したブレブの根元部分には、特徴的な膜構造が形成されている様子が観察されます。
下段左は、ブレブの根元部分を三次元電子顕微鏡解析(CLEM)により詳細に観察した結果です。細胞外空間を緑色、ブレブ基部に存在する膜構造を赤色で示しています。観察の結果、ブレブの根元には細胞膜から連続した細長い膜の陥入構造が存在することが明らかになりました。下段右は、その構造の模式図です。本研究では、この新たに見出された膜構造を「Sub-bleb invagination; SBI」と名付けました。SBIは、ブレブが拡大する際にその場で形成される特徴的な膜構造であり、膜タンパク質の配置を制御する役割を持つと考えられます。
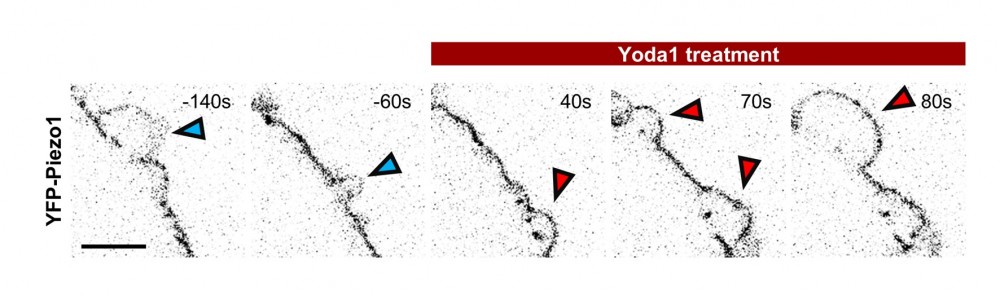
YFPで標識したPiezo1の細胞内分布をライブセルイメージングにより観察したものです。画像は反転表示で示しています。Yoda1添加前では、Piezo1は主にブレブの根元に形成されるSBIに集積し、外側に突出したブレブ膜にはほとんど局在しません(青矢印)。一方、Piezo1の構造を変化させる活性化化合物であるYoda1(50 µM)を添加すると、Piezo1の構造が変化し、膜の曲がり方に対する選好性(膜曲率選好性)が変わります。その結果、Piezo1はSBIだけでなく、外側に突出したブレブ膜にも分布するようになります(赤矢印)。
この結果は、膜タンパク質の局在がその膜曲率選好性によって決定されること、そしてその性質を変化させることで細胞膜上での配置が動的に変わることを示しています。
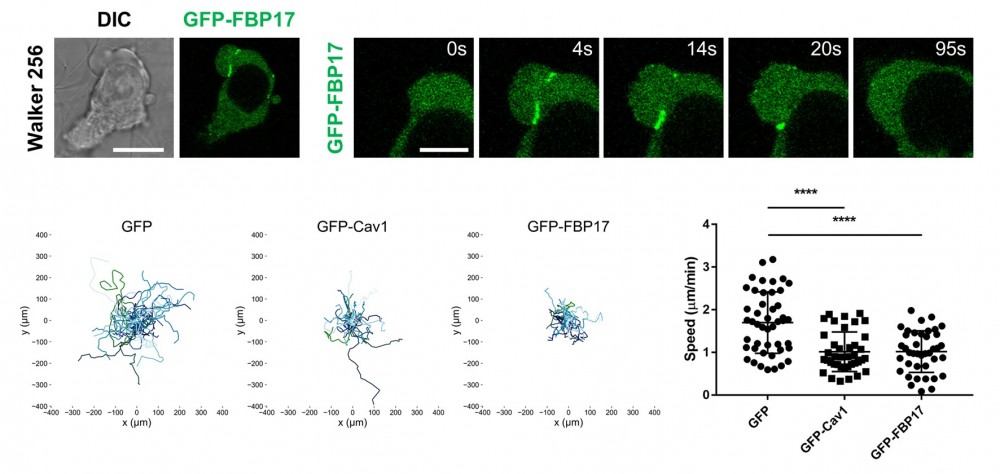
上段では、活発にブレブを形成し高い遊走性を示すがん細胞Walker256細胞において、SBIが形成されている様子を観察しています。SBIに集積する分子であるFBP17にGFPを付加したタンパク質を発現させることで、SBIの形成を可視化しています。
下段左では、Walker256細胞において、内向きの膜曲率を好むタンパク質であるCaveolin-1およびFBP17を過剰に発現させると、膜の内向き陥入構造が過剰に形成され、細胞の移動が抑制されることを示しています。下段右では、GFP(対照)、GFP-Caveolin-1、GFP-FBP17をそれぞれ発現させた細胞の移動速度を定量しています。その結果、Caveolin-1およびFBP17を発現させた細胞では、対照細胞と比べて移動速度が有意に低下していました。
これらの結果は、内向きの膜曲率を好むタンパク質が増加すると膜の折れ込み構造が過剰に形成され、その結果としてブレブ形成および細胞運動が抑制されることを示しています。すなわち、膜構造の制御が細胞の移動能力を規定することが明らかになりました。
(※1) Piezo1
細胞膜に存在し、膜の変形や力(機械刺激)を感知するタンパク質(機械受容チャネル)です。細胞膜が押されたり引き伸ばされたりすると開いて、カルシウムイオンなどを細胞内に取り込み、外からの刺激を細胞内に伝える働きを持ちます。Piezo1は膜の「曲がり方」に対する好み(膜曲率選好性)を持っており、特定の形状の膜に集まりやすい性質があります。本研究では、この性質が変化すると、Piezo1が細胞膜上のどこに分布するかが変わることが示されました。
(※2) ブレブ
細胞膜が一時的に外側へ膨らむ現象です。アポトーシス(細胞死)や細胞分裂の際に受動的に形成されるだけでなく、がん細胞や免疫細胞では、細胞が移動するために能動的に形成されます。このようなブレブを利用した運動は、組織内での移動や浸潤に重要な役割を果たします。
- 掲載誌:
- Proceedings of the National Academy of Sciences of the United States of America(PNAS)
- タイトル:
- Bleb expansion requires transient membrane invaginations that sequester curvature-preferring proteins
- 著者名:
- 前川悠輝、吉井紗織、水島昇、池ノ内順一
- DOI:
- 10.1073/pnas.2534871123
医学研究院 生化学分野
教授 池ノ内 順一(イケノウチ ジュンイチ)
TEL:092-642-6096 FAX:092-642-6096Mail:ikenouchi.junichi.033(at)m.kyushu-u.ac.jp
