2021.05.19
「レット症候群原因因子による神経幹細胞の分化制御メカニズムを明らかに」(基盤幹細胞学分野 中島 欽一教授)
レット症候群原因因子による神経幹細胞の分化制御メカニズムを明らかに
〜発達障害の発症メカニズムの解明と新たな治療法開発に期待〜
〜発達障害の発症メカニズムの解明と新たな治療法開発に期待〜
概 要
九州大学大学院医学研究院の中嶋秀行助教、中島欽一教授らの研究グループは、広島大学大学院統合生命科学研究科の今村拓也教授、名古屋大学大学院理学研究科・高等研究院の辻村啓太特任講師、慶應義塾大学医学部生理学教室の岡野栄之教授らとの共同研究により、神経発達障害レット症候群(※1)の原因因子である methyl-CpG binding protein 2(MeCP2)(※2)がマイクロRNA(miRNA)(※3)を介して神経幹細胞(※4)の分化を制御していることを発見し、そのメカニズムを明らかにしました。
レット症候群は自閉症、てんかん、失調性歩行、特有の常同運動(手もみ動作)を主徴とする進行性の神経発達障害です。MeCP2 遺伝子の変異により発症することはわかっているもののその発症機序の詳細は不明でした。本研究グループは脳の発生過程において、MeCP2 が神経幹細胞のニューロンへの分化を促進し、通常はニューロンの機能を支持するアストロサイトへの分化は抑制していることを明らかにしました。また、そのメカニズムについて調べた結果、MeCP2 は miR-199aという miRNA を介して脳の発達に重要な骨形成因子(BMP)シグナル(※5)を抑制することで、神経幹細胞の分化を制御していることがわかりました。さらに、MeCP2 遺伝子に変異をもつレット症候群患者由来の iPS 細胞から作製した脳オルガノイド(※6)では BMP シグナルの亢進とアストロサイトへの分化増加がみられ、これらが BMP シグナル阻害剤により改善できることが明らかになりました。以上の結果は、レット症候群患者脳では、神経幹細胞からニューロンやアストロサイトへの分化バランスが上手く制御されていない可能性を示しており、そのバランスの正常化によるレット症候群の新しい治療法開発へとつながることが期待されます。
本研究成果は、2021 年 5 月 18 日(火)午前 11 時(米国東部標準時間)に国際学術雑誌『Cell Reports』に掲載されました。なお、本研究は文部科学省科研費、日本医療研究開発機構(AMED)、精神・神経疾患研究開発費、レット症候群支援機構の支援を受けました。
九州大学大学院医学研究院の中嶋秀行助教、中島欽一教授らの研究グループは、広島大学大学院統合生命科学研究科の今村拓也教授、名古屋大学大学院理学研究科・高等研究院の辻村啓太特任講師、慶應義塾大学医学部生理学教室の岡野栄之教授らとの共同研究により、神経発達障害レット症候群(※1)の原因因子である methyl-CpG binding protein 2(MeCP2)(※2)がマイクロRNA(miRNA)(※3)を介して神経幹細胞(※4)の分化を制御していることを発見し、そのメカニズムを明らかにしました。
レット症候群は自閉症、てんかん、失調性歩行、特有の常同運動(手もみ動作)を主徴とする進行性の神経発達障害です。MeCP2 遺伝子の変異により発症することはわかっているもののその発症機序の詳細は不明でした。本研究グループは脳の発生過程において、MeCP2 が神経幹細胞のニューロンへの分化を促進し、通常はニューロンの機能を支持するアストロサイトへの分化は抑制していることを明らかにしました。また、そのメカニズムについて調べた結果、MeCP2 は miR-199aという miRNA を介して脳の発達に重要な骨形成因子(BMP)シグナル(※5)を抑制することで、神経幹細胞の分化を制御していることがわかりました。さらに、MeCP2 遺伝子に変異をもつレット症候群患者由来の iPS 細胞から作製した脳オルガノイド(※6)では BMP シグナルの亢進とアストロサイトへの分化増加がみられ、これらが BMP シグナル阻害剤により改善できることが明らかになりました。以上の結果は、レット症候群患者脳では、神経幹細胞からニューロンやアストロサイトへの分化バランスが上手く制御されていない可能性を示しており、そのバランスの正常化によるレット症候群の新しい治療法開発へとつながることが期待されます。
本研究成果は、2021 年 5 月 18 日(火)午前 11 時(米国東部標準時間)に国際学術雑誌『Cell Reports』に掲載されました。なお、本研究は文部科学省科研費、日本医療研究開発機構(AMED)、精神・神経疾患研究開発費、レット症候群支援機構の支援を受けました。
 |
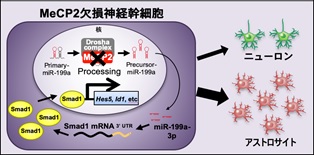 |
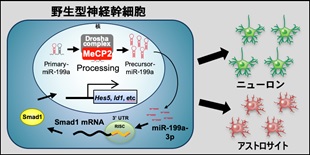
| (参考図)MeCP2 は神経幹細胞の分化バランスを制御する 野生型神経幹細胞では、MeCP2 と Drosha 複合体が Pri-miR-199a をプロセシングし、正常なレベルのMature-miR199a-3p が産生される。Mature-miR199a-3p は BMP 下流転写因子 Smad1 を標的とすることでニューロン分化とアストロサイト分化の適度なバランスを制御する(左図)。 一方、MeCP2 欠損神経幹細胞では、MeCP2 欠損のため Drosha 複合体が Pri-miR199a を正常にプロセシングできず、Mature-miR199a-3p が十分量産生されない。その結果、Smad1 タンパク質の異常増加により、ニューロン分化が抑制され、アストロサイト分化優位へとバランスがシフトする(右図)。 |
|
| 【お問い合わせ】 | 九州大学大学院医学研究院 基盤幹細胞学分野 教授 中島 欽一 | |
| TEL:092-642-6195 FAX:092-642-6561 Mail:nakashima.kinichi.718(a)m.kyushu-u.ac.jp ※(a)を@に置き換えてメールをご送信ください |
■背 景
X 染色体上に存在する methyl-CpG binding protein 2(MeCP2)遺伝子の変異は、レット症候群をはじめ、自閉症、てんかん、統合失調症などを含めた様々な精神疾患・神経発達障害への関与が示唆されています。レット症候群は獲得された運動、言語能力の喪失、精神遅滞、自閉症傾向などを示す進行性の神経発達障害であり、神経系細胞特異的にMeCP2 を欠損したマウスはレット症候群患者と類似の表現型を示すことから、MeCP2 の神経系での機能が重要であることが示唆されているものの、レット症候群の表現型に関与する決定的な下流標的因子は未だに同定されておらず、レット症候群発症機序の全貌は不明なままでした。
我々の研究グループは以前、MeCP2 が microRNA (miRNA) マイクロプロセッサーである Drosha 複合体と会合し、特定の miRNA(miR-199a)のプロセシングを促進することを見出していました。また、レット症候群の死後脳では miR-199a の発現が減少しており、miR-199a 欠損マウスは MeCP2 欠損マウスと同様の表現型を示すことも明らかにしていました。しかし、以前の研究は、神経幹細胞から産生されたニューロンでの MeCP2 の機能にのみ焦点を当てたもので、ニューロン以外にもアストロサイトなど、他の神経系細胞を生み出し、脳発生に重要な神経幹細胞における機能は不明のままでした。そこで、脳の発達過程における MeCP2 及び miR-199a の機能を明らかにすることで、MeCP2 遺伝子の機能異常が原因で引き起こされる発達障害のさらなる発症メカニズムに迫ろうと考えました。
■内 容
研究グループは生後 1 日目の野生型及び MeCP2 欠損、miR-199a 欠損マウス脳から海馬を取り出し 1 細胞遺伝子発現解析を行いました。その結果、MeCP2 欠損及び miR-199a 欠損海馬では共通してニューロン細胞集団が野生型海馬より減少している一方、アスロトサイト細胞集団は増加していることが明らかとなりました。この結果より、MeCP2 欠損及び miR-199a 欠損マウス脳では神経幹細胞の分化バランスがニューロンからアストロサイトへシフトしている可能性が考えられました。そこで、胎生 14 日目の野生型及び MeCP2 欠損、miR-199a 欠損マウスから神経幹細胞を単離し、培養皿の上で分化誘導を行った結果、MeCP2 欠損及び miR-199a 欠損神経幹細胞はニューロンへの分化が減少し、アストロサイトへの分化が増加していることが確かめられました。また詳細な解析により、miR-199a は脳の発達過程に重要な機能をもつ BMP シグナルの下流転写因子 Smad1 をターゲットにすることで神経幹細胞の分化を制御していることを突き止めました。
次に、これまでマウスを用いて得られた結果と同様の現象が、実際のレット症候群患者の脳においても引き起こされている可能性を調べる目的で、レット症候群患者由来の iPS 細胞から脳オルガノイドを作製し検討を加えました。その結果、レット症候群患者 iPS 細胞由来脳オルガノイドではアストロサイトへの分化が亢進しており、Smad1 のタンパク質量も増加していることがわかりました。さらに、レット症候群患者 iPS 細胞由来脳オルガノイドへ BMP シグナル阻害剤を添加することで、これらの表現型を改善できることを見出しました(図 1)。
このような結果から、MeCP2 は miR-199a を介して神経幹細胞の分化運命決定を制御しており、MeCP2/miR-199a/Smad という分子ネットワークがレット症候群病態において重要な役割を果たしていることが明らかになりましたた。
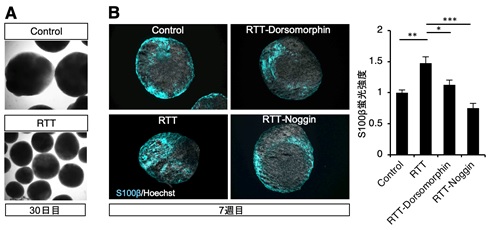
図 1: 培養 30 日目の正常(Contorl)及び変異 MeCP2(RTT)を発現する脳オルガノイドの写真(A)。BMP シグナル阻害剤(Dorsomorphin,Noggin)を添加するとアストロサイトマーカー(S100β)の蛍光強度が減少する(B)。
■効果・今後の展開
今回の研究結果により、レット症候群の原因因子である MeCP2 が脳の発達過程において重要な役割を担っていることがわかると同時に、MeCP2 変異を持つレット症候群の病態に新たな知見を提供しました。今回の成果を基に、今後は、BMP シグナル阻害による方法なども含めて、レット症候群を含めた発達障害の新規治療法開発につながることを期待しています。
【用語解説】
(※1)レット症候群
自閉症やてんかん、失調性歩行、特有の常同運動(手もみ動作)を主徴とする進行性の神経発達障害である。X連鎖優性遺伝病であり、男性は胎生致死で女性のみが罹患する。レット症候群の80-90%にMeCP2遺伝子の変異がみられる。
(※2)MeCP2
MeCP2の遺伝子はX染色体上に存在。MeCP2はメチル化された遺伝子のプロモーター領域に結合し、標的遺伝子の発現を抑制する転写抑制因子として同定されたが、私達の以前の研究によって、今回の研究でも重要な役割を果たすmiRNAの生合成にも関わることが明らかとなっている。MeCP2遺伝子の変異は、レット症候群の原因となるだけでなく、自閉症、双極性障害、認知障害、統合失調症患者にも認められる。
(※3)マイクロRNA(miRNA)
細胞内に存在する長さ18から24塩基程度の1本鎖RNAであり、遺伝子の転写後発現調節に関与している。数百から数千塩基の一次前駆体(Primary-RNA)として転写され、核内でDrosha複合体にプロセシングされる。
(※4)神経幹細胞
自己複製能および脳を構成する主要な細胞種であるニューロン、アストロサイト、オリゴデンドロサイトへの多分化能を併せもつ幹細胞。
(※5)BMPシグナル
BMPは、セリン/スレオニンキナ-ゼドメイン型受容体を介して、転写因子Smad1を含む下流因子を活性化することで細胞内にシグナルを伝達する。
(※6)脳オルガノイド
多能性幹細胞(iPS細胞やES細胞)などを用いて、脳の形成過程を体外で模倣し、作製される三次元組織。
| 論文名: | MeCP2 controls neural stem cell fate specification through miR-199a-mediated inhibition of BMP-Smad signaling. |
| 著者名: | Hideyuki Nakashima, Keita Tsujimura*, Koichiro Irie, Takuya Imamura, Cleber A. Trujillo, Masataka Ishizu, Masahiro Uesaka, Miao Pan, Hirofumi Noguchi, Kanako Okada, Kei Aoyagi, Tomoko Andoh-Noda, Hideyuki Okano, Alysson R. Muotri, and Kinichi Nakashima* (*共責任著者) |
| 掲載誌: | Cell Reports, 2021 |
| DOI: | 10.1016/j.celrep.2021.109124 |
【謝辞】
本研究は文部科学省科研費(JP17H01390, JP16H06527, JP16K21734, JP16H06279 (PAGS), JP16J03827 and JP19K16918)、日本医療研究開発機構(AMED) (JP20mg1310008)、精神・神経疾患研究開発費(27-7 and 30-9)、レット症候群支援機構(2019-01-09,10)の支援を受けました。
