2021.06.23
「発達期に特定の樹状突起が選択的に強化される仕組みを解明」(疾患情報研究分野 今井猛教授)
発達期に特定の樹状突起が選択的に強化される仕組みを解明
~分泌タンパク質BMPと神経伝達物質の共入力が鍵~
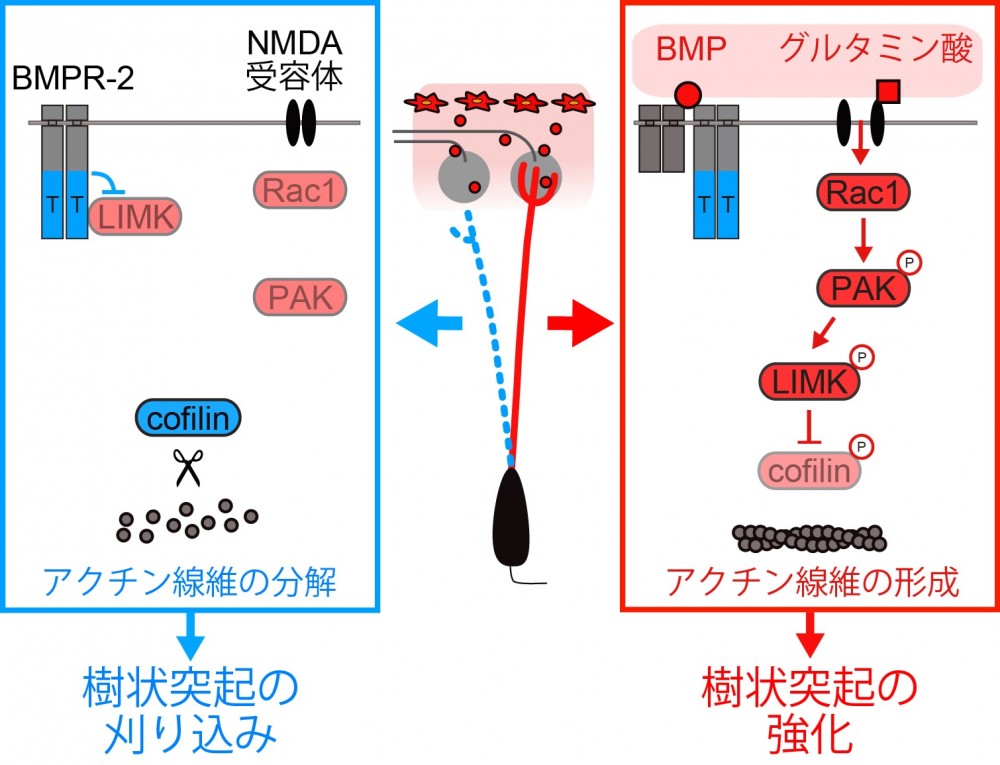
脳の神経回路を構成する神経細胞は、樹状突起と呼ばれる神経突起を介して情報を入力します。神経細胞に正しい情報のみを入力するには、発達期に樹状突起が正しく配線される必要があります。匂いの情報処理に関わる嗅球の僧帽細胞と呼ばれる神経細胞は、出生直後までに樹状突起を複数伸ばします。その後、生後発達期にはそのうちの1つだけを強化し、その他を刈り込むことで正しい配線を獲得します。しかしながら、僧房細胞がどのようにして正しい接続とそれ以外を区別し、樹状突起の選択的な強化・刈り込みを行っているのかはわかっていませんでした。本研究では、BMPR-2と呼ばれる細胞表面分子が樹状突起の選択的強化・刈り込みの鍵となっていることを明らかにしました。BMPR-2は、BMPという分泌タンパク質があるときには神経伝達物質グルタミン酸のシグナルが入力されるようにし、樹状突起内のアクチン細胞骨格系の強化を促します。これにより、樹状突起は安定化され、シナプスの形成が促進されます。一方で、BMPがないときにはグルタミン酸による入力の情報が遮断され、下流に伝わらないため、逆に樹状突起の刈り込みが促進されます。ひとたび樹状突起が強化されると、強化するためのシグナルがますますたくさん入るようになるため、より強固な樹状突起が作られます。
本研究では嗅球の僧帽細胞をモデルとして研究を行いましたが、BMPR-2は大脳皮質におけるシナプス形成や、神経発達障害のひとつである脆弱X症候群との関連も指摘されています。今回の発見が神経発達障害の発症メカニズムの理解に貢献することが期待されます。
本研究は JST さきがけ、CREST、新学術領域研究「スクラップ&ビルドによる脳機能の動的制御」、日本学術振興会(JSPS)科学研究費補助金、持田記念医学薬学振興財団、上原記念生命科学財団の助成によって行われました。本成果は、令和 3 年 6 月 22 日(火)(米国時間)に米国のオンライン科学雑誌『Cell Reports』に掲載され、表紙を飾りました。本研究は九州大学および理化学研究所多細胞システム形成研究センター(当時)にて実施されました。
|
研究者からひとこと:発達期の神経細胞は、自発的に正しい標的を判断してつながりを作ります。ここで間違ったつながりを作ってしまうと精神疾患の原因にもなります。この不思議で重要な謎を解き明かすために日々研究をしています。
|
 |
 |
|
|
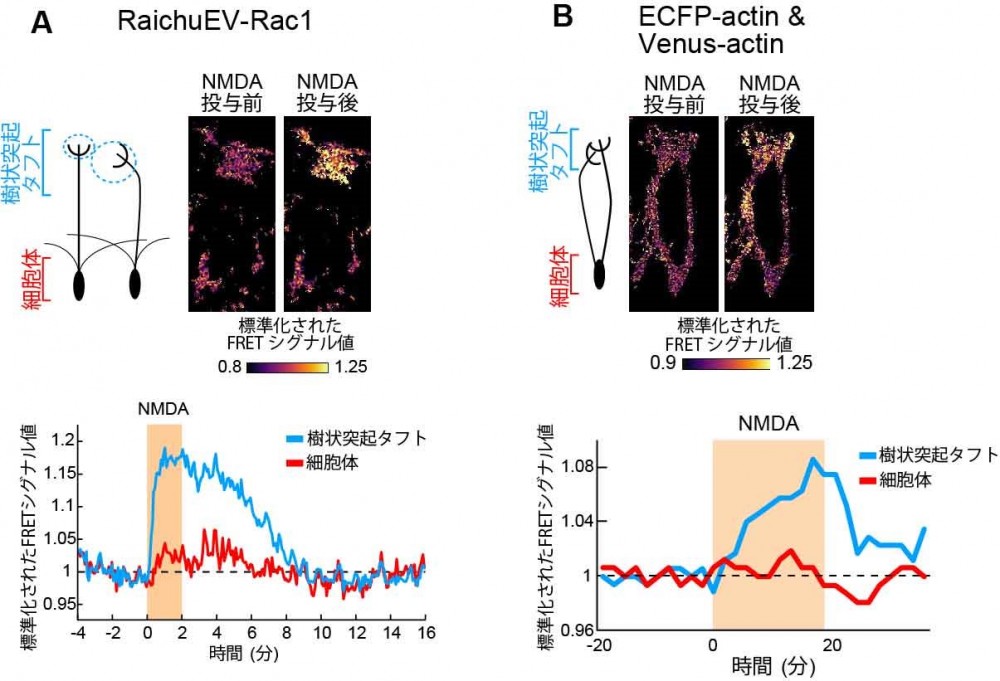
(図1)BMPという分泌タンパク質と神経伝達物質グルタミン酸の入力が両方ある時のみ、細胞内で細胞の骨組みであるアクチン線維が形成され、樹状突起が強化される(右)。逆に入力が無いときはアクチン線維が分解され樹状突起の刈り込みにつながる(左)。
|
|
| 【お問い合わせ】 | 大学院医学研究院 教授 今井 猛 Mail: imai.takeshi.457(a)m.kyushu-u.ac.jp | |
| ※(a)を@に置きかえてメールをご送信ください。 |
動物は感覚器官(目や鼻など)で受け取った外界の情報を神経細胞において電気信号へと変換します。この電気信号は神経伝達物質を介して脳内の神経細胞へと次々に伝達されます。脳内の神経細胞は樹状突起と呼ばれる突起から情報を入力し、軸索と呼ばれる突起を用いて情報を出力します。樹状突起と軸索を介して情報が神経細胞間を次々とリレーされていくことで、脳は外界の情報を知覚し、行動へとつなげることができます。適切な情報処理を行う上では、樹状突起と軸索の適切な配線が重要です。例えば匂い情報を司る脳部位、嗅球に存在する僧房細胞と呼ばれる神経細胞は、主樹状突起と呼ばれる太い樹状突起を1つだけ伸ばし、1種類の嗅覚受容体(匂いセンサー)の情報のみを入力しています。ここで、もし複数の主樹状突起がつくられてしまうと、僧帽細胞には複数の匂いの情報が入力してしまうこととなり、情報が混線してしまうことになります(図2)。従って、僧帽細胞においては、主樹状突起を1つだけ配線することが重要です。
■研究手法と成果
■今後の期待
生後発達期において特定の樹状突起を強化し、それ以外の樹状突起を刈り込む「再編成」という現象は脳内の多くの神経細胞で知られていましたが、その仕組みはわかっていませんでした。本研究では、グルタミン酸入力による樹状突起強化の「アクセル」シグナルに対し、BMPR-2がクラッチとしての役割を果たし、両者の共入力によって初めて樹状突起の強化が生じることが明らかになりました。このように複数のシグナルの共入力によって樹状突起が強化されることは、適切な配線を保証するうえで普遍的な仕組みである可能性があります。今後、本研究結果を足掛かりに、他の神経細胞でも同様の仕組みが発見されることが期待されます。
■本研究について
本研究は、JSTさきがけ、CREST、新学術領域研究「スクラップ&ビルドによる脳機能の動的制御」(JP16H06456)、日本学術振興会(JSPS)科学研究費補助金(JP23680038, JP15H05572, JP15K14336, JP16K14568, JP16H06456, JP17H0626, JP15K18353)、持田記念医学薬学振興財団、上原記念生命科学財団の助成によって行われました。また、本研究は九州大学および理化学研究所多細胞システム形成研究センター(当時)にて実施されました。
■用語解説
(※1)CRISPR/Cas9:
元々は細菌の免疫機構として発見された。細菌はウイルスが侵入してきた時に、ウイルス遺伝子の一部を自身のゲノム(CRISPR領域)に保存する。2回目のウイルス侵入時は保存されたウイルス遺伝子の配列を使うことで、ウイルス遺伝子を外部からの侵入者として認識。Cas9タンパクによりそのウイルス遺伝子を切断することで、ウイルスの増殖を防ぐ。狙った遺伝子を特異的に切断する能力を利用して、近年遺伝子編集ツールとして利用されている。この技術を開発したシャルパンティエ博士とダウドナ博士は2020年にノーベル化学賞を受賞した。
(※2) 子宮内電気穿孔法:
脳に存在する神経幹細胞に遺伝子を導入する技術。特定の神経細胞に対して、容易に遺伝子操作を行うことができる。
(※3) 神経幹細胞:
神経細胞の元となる細胞。多くの神経幹細胞は胎児期に分裂し、神経細胞を生み出す。
(※4) 分泌タンパクBMP:
細胞内で産生され、細胞外に分泌されるタンパクを分泌タンパク質と呼ぶ。BMP(Bone Morphogenetic Protein;骨形成タンパク質)は、当初骨の形成を促進するタンパク質として発見されたが、その後、胎生期の体軸形成や細胞運命決定など、様々な機能を有していることが後にわかった。
(※5) アクチン線維:
細胞骨格の1種。アクチン線維とは単量体の球形アクチンがいくつもつながってできた線維状のものを指す。
(※6) 遺伝子操作:
外部からDNAや遺伝子などを細胞に導入することで、ゲノム配列やタンパク質の発現を操作すること。CRISPR/Cas9も含まれる。本研究ではCRISPR/Cas9による遺伝子ノックアウトに加え、特定遺伝子(LIMK1など;図4~6参照)を子宮内電気穿孔法により細胞内に導入し、そのタンパク質を発現させた。
(※7) FRETイメージング:
FRET(Förster Resonance Energy Transfer)、日本語でフェルスター共鳴エネルギー移動と呼ばれる。特定の2種類の蛍光分子が近傍にある状態で、短波長側の蛍光分子の励起光を当てると、励起されたエネルギーが長波長側の蛍光分子に移動し、長波長側の励起光が放出される現象のこと。これを利用したセンサーをFRETセンサー、イメージング手法をFRETイメージングと呼ぶ。一般的に、タンパク質の活性変化には構造変化を伴う。従って、この時に構造的に近づいたり離れたりする位置に蛍光タンパク質を結合させておくと、活性の変化をFRETシグナルとして検出することができる。本研究ではRac1の活性化やアクチン線維形成を計測するために利用。
(※8) シナプス:
| タイトル: | BMPR-2 gates activity-dependent stabilization of primary dendrites during mitral cell remodeling (BMPR-2は僧房細胞のリモデリングにおける神経活動依存的な樹状突起安定化を制御する) |
| 著者: | 藍原周平、藤本聡志、坂口理智、今井猛 |
| 掲載誌: | Cell Reports |
| DOI: | https://doi.org/10.1016/j.celrep.2021.109276 |
■参考図
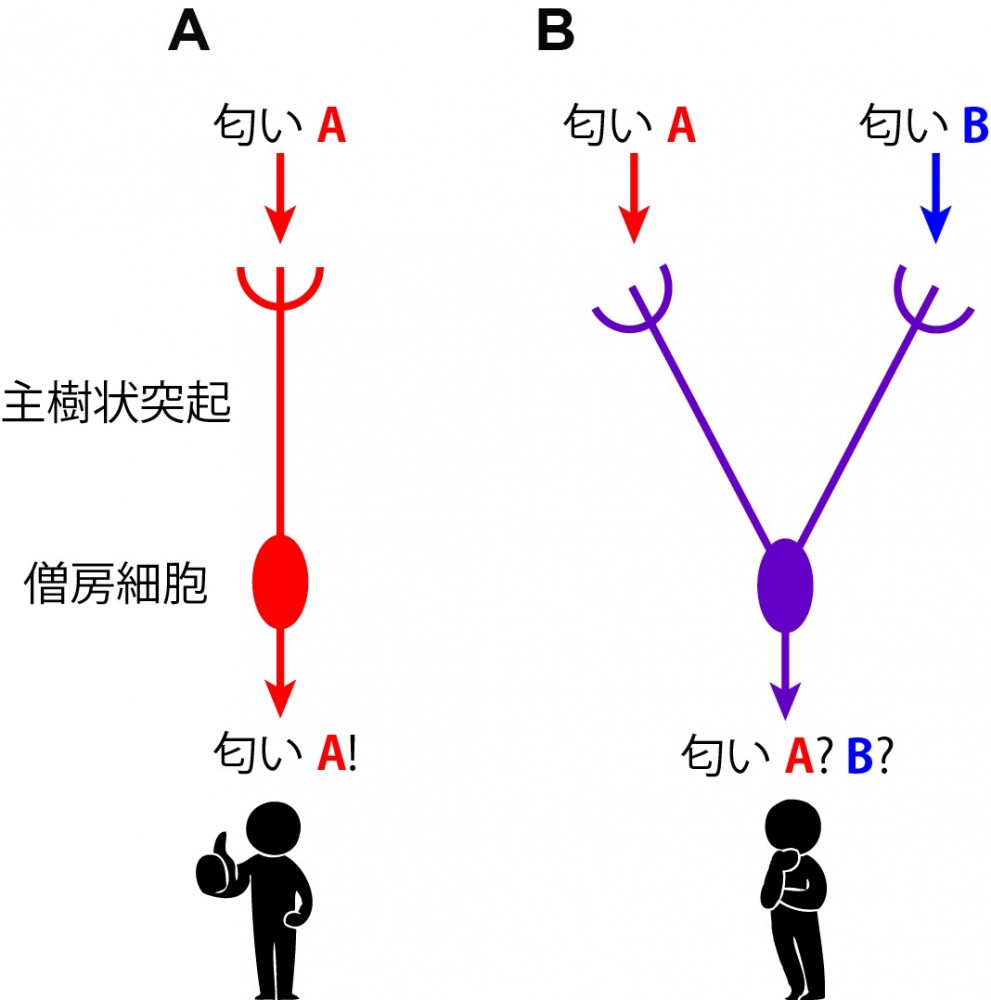 |
(図2) 僧房細胞の主樹状突起を介した匂い情報入力 (A)正常な僧房細胞は1つの主樹状突起を介して、1種類の嗅覚受容体(匂いセンサー)の情報のみを受け取る。これが匂い識別の基礎となっている。(B)もし、僧帽細胞が間違って複数の主樹状突起を形成してしまうと、1つの僧房細胞に複数の匂い情報が入力されてしまうため、匂いの識別ができなくなってしまう。このように樹状突起の正しい配線は感覚情報の精緻な識別に必要である。 |
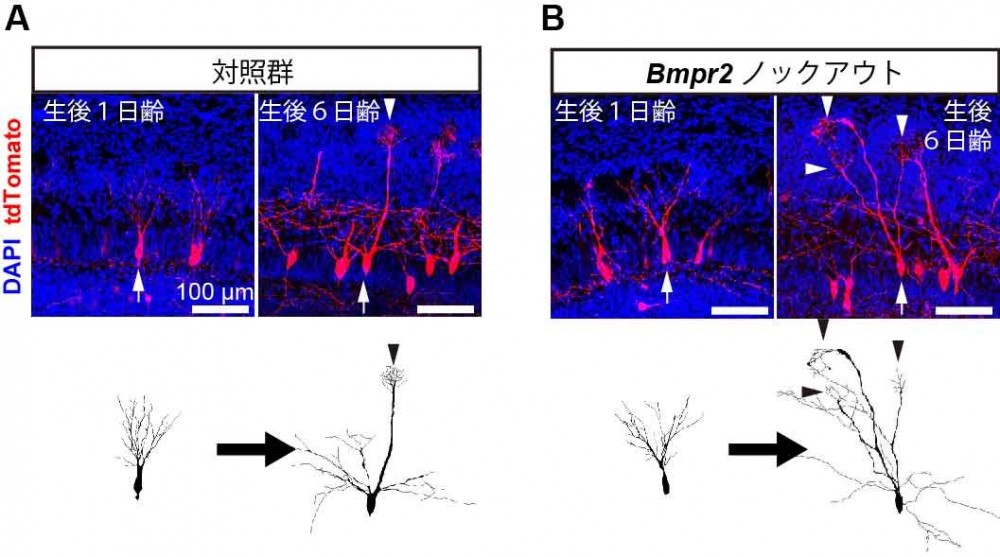
(A)通常、僧房細胞は複数の樹状突起を嗅球表面(写真上方向)に伸ばすが、生後6日齢までには1つの主樹状突起(矢頭)のみを持つようになる。(B)CRISPR/Cas9を用いてBMPR-2遺伝子を欠損させると、成長後も複数の主樹状突起をもつ僧房細胞が多くみられるようになる。個々の僧房細胞の形態を抽出して下に示す。
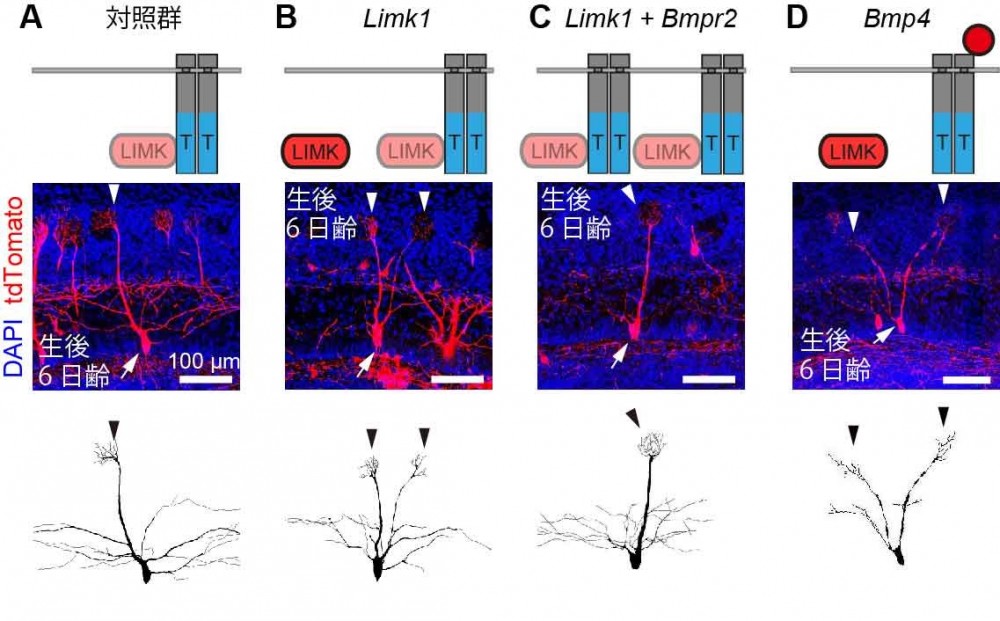
(図4) BMPR-2はBMPがない状態ではLIMKを抑制する
(A)生後6日齢の正常な僧房細胞。ほとんどの樹状突起でBMPR-2がLIMKを抑制するため、主樹状突起は1つしか形成されない。(B)LIMK1を過剰発現させた僧房細胞。BMPR-2で過剰量のLIMKを抑制しきれず、複数の主樹状突起ができてしまう。(C)LIMK1に加えてBMPR-2を過剰発現させた僧房細胞。過剰量のLIMKは過剰量のBMPR-2により抑制されるため、主樹状突起は1つだけ生じる。(D)BMPの1種であるBMP4を過剰発現させた僧房細胞。全ての樹状突起で分泌されたBMP4がBMPR-2に結合するため、LIMKの抑制が解除され、複数の主樹状突起が形成される。